In my last week this summer at MBL, I decided to do a biological experiment. And why not? Well why not is because to do the experiment correctly, I need the three-dimensional solution that is still on the coding board. But let’s not let perfect be the enemy of done. Even if the experiment would not be strictly quantitative, I thought I might as well have a look. You may look too (figures below) but first here is what I did.
I transferred regular BY-2 cells (not the xylem line) into two 15 mL flasks and in one flask I added an inhibitor of microtubule polymerization (a compound called propyzamide, which is widely used for this purpose in BY-2 cells). Without microtubules, cellulose is laid down in arbitrary directions rather than perpendicular to the long axis of the cell. Without uniform cellulose, the cell swells and after a few days becomes spherical. But how, exactly, does the cellulose lose its organization? And what is the relation between changed cellulose and the changed growth pattern? I have been looking at this in roots but roots have thousands of cells. I thought that looking at this single cell system would provide a valuable comparison. I still do.
About 24 h after adding the drug, I took half a milliliter of cell suspension from each culture to have a look. To make absolutely sure that sampling the culture on day one did not contaminate it for day two, I took the trouble of sampling the cultures in the sterile hood upstairs. This is a bit of a faff because I have to schlep the gear I need up there but worth it withal. Because I planned to observe polarized fluorescence, I had a pair of tubes with half a mL of fast scarlet, each, ready and waiting beside the hood so I could add the cells directly. The cells bathe in stain for half an hour and are then rinsed. To rinse them I use an old, hand-crank centrifuge, which is clamped onto the bench right next to the tunicate tank. Its thin and rusty arms partially resemble a dubious umbrella as they open out on hinges while the baskets spin. The bench shudders and I worry about something coming loose, but I only have to count to 12 to spin the cells down. Then, I remove the solution containing the dye above the pellet of cells, add 3% sucrose and repeat. After that, the cells are ready to be observed. I observed cells on day 1, day 2, and day 3.
I placed 20 µL of the rinsed suspension of cells on a glass slide and add a coverslip. Now, I could observe. But, the slide will be under the microscope for an hour or more, and water would evaporate out from the edges of the coverslip letting in air, which ruins the image (and isn’t so nice for the cells). The coverslip has to be sealed. Rudolf has nifty slide-sealing gear. He took a power supply for a typical microscope light, probably 12 V DC, and attached a household electrical plug to the output leads. Between the leads, he ran a thick wire, molding in into a long narrow loop. When the power is switched on, this gets hot. There is a pot of wax (I believe a mixture of Vaseline, lanolin, and paraffin – yellow and waxy) and holding the plug by the safe part (relatively safe part!), I poke the narrow wire loop gently into the wax, thereby transferring a little bit of the wax onto the loop where it melts, but the liquid is held in the space between the wires. Chopsticks anyone? I then touch down this drop of molten wax along the edge of the coverslip, forming a bead. This sets in a few seconds. With a few such transfers, the whole coverslip is sealed to the slide.
Then onwards to the microscope. I took images in retardance mode, where the organization state of the cellulose is imaged directly, and for the same cells, images of polarized fluorescence from fast scarlet. I used both modes because they differ. The retardance mode provides information for the entire cell wall thickness whereas the dye reports mostly the surface layer (probably both inner and outer surfaces, but not throughout the wall’s thickness. On the other hand, retardance will be directly compromised by the other wall of the cell, that is the one across the vacuole and cytoplasm on the other side. It is precisely to avoid this interference that I want 3-D power. That other wall will affect retardance more than fluorescence because for retardance the beam goes thru both walls whereas for fluorescence it is only the overlay of out of focus fluorescence that interferes. Because the other side of the wall is 30 µm distant, the out-0f-focus fluorescence will probably be pretty weak.

Well, the drug does a number on the wall. The rather uniform deployment of cellulose is turned topsy-turvy (Fig. 1, 2). The images from retardance and polarized fluorescence are qualitatively similar, but differ in detail. I am not sure how I can quantify this or even if I will. That wiping out the microtubules causes cellulose deposition to go nuts is really no surprise. It would be interesting to see quantitatively how close (or not) the two imaging modes agree. The effects of the drug were less severe on day 1 and that might be worth quantifying.
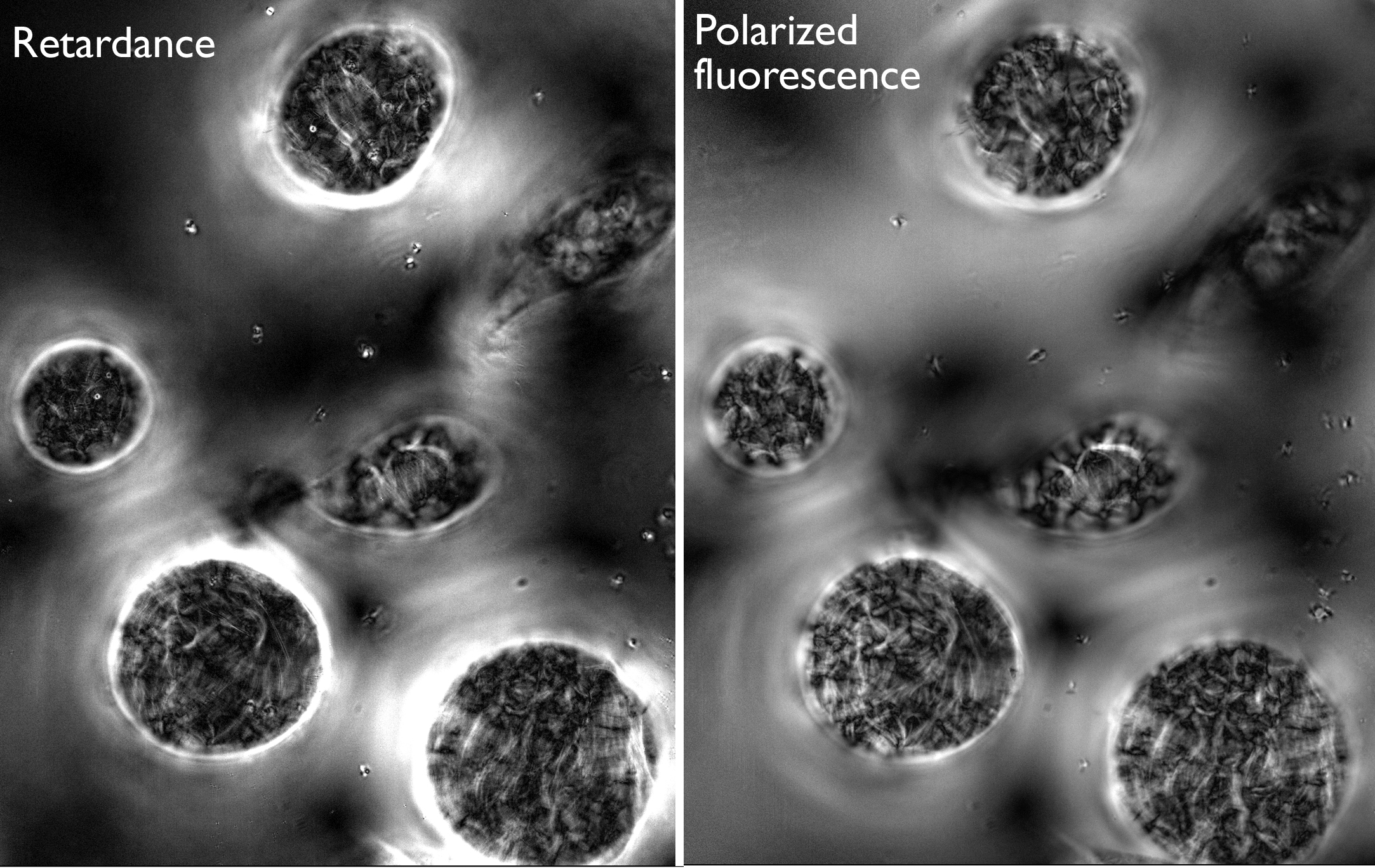
But the key experiment, for the future, will be to use a lower level of drug, one that causes some cell swelling but does not turn the cells into beach balls. There will be some microtubules present, exerting some influence over the cellulose–but what exactly? I just might have to figure out how to do this at home!